 
 The LUHNA Book!
|

  |
|
by
Mark A. Withers
Michael W. Palmer
Gary L. Wade
Peter S. White
Paul R. Neal
Abstract. In this study we used data from published floras to produce contour maps of flora size and percentage of exotic species in North America for two specified unit areas (1,000 and 1,000,000 ha) and two specified years (1900 and 1996). Flora size decreases toward higher latitudes and is greatest in the southeastern United States. The number of species in floras decreases from east to west. This decrease may result from drier environments, which have been linked to reduced diversity, and to a less thoroughly documented flora. Increases in the size of floras since 1900 probably do not reflect increased plant biodiversity but rather suggest that plant life has been more thoroughly documented. The average size of floras in parts of the eastern United States has decreased since 1900, but this decrease may be a result of increased botanical interest in unusual, but relatively depauperate, environments. Proportions of exotic species are greatest in the northeastern United States. The west coast also has relatively higher proportions of exotic species. The proportion of exotic species is least in the mountain West and in the far northern latitudes of Canada and Alaska. Despite the lack of complete or consistent scientific data, we believe that floras represent a valuable data source to assess biodiversity.
Biodiversity and Exotic Species In recent years, scientists, government officials, and the general public have become increasingly concerned over growing threats to biological diversity. Biological diversity--or biodiversity--represents the variation in life forms that exist on earth. This variation occurs at every level of the taxonomic hierarchy, but most discussion of biodiversity focuses on species diversity. Every species on earth plays a unique role in maintaining and balancing ecosystem processes and functions. The high level of interdependence of species in ecosystems suggests that a reduction of biodiversity at any taxonomic level could produce undesirable results. For example, virtually all animal life depends directly on the food energy trapped by plants through photosynthesis. However, plant life also depends on other organisms, such as microorganisms and small animals (macroinvertebrates) that return to the soil the nutrients that plants need. Similarly, plants often depend on insects or birds for pollination and dispersal. Reduced biodiversity of any of these groups would certainly affect the other groups (Wilson 1988). Human life is also bound to the diversity of plants and animals (as well as other types of organisms such as fungi and bacteria). Although all groups of living organisms are vitally important to humans, we focus here on plants. Plants provide not only food energy but also a host of other products, such as rubber, fiber, medicines, and fuel. In recent years, plants have yielded valuable drugs to treat malaria, heart disease, various cancers, and other ailments, yet only a tiny fraction of the world's plant species has been studied in detail. Pharmacologists believe many thousands of other useful medications remain undiscovered in the world's plants. Plants are also vital to maintaining and improving the overall health and function of the world's ecological systems. These "ecosystem services" include regulation of rainfall and runoff, erosion control, flood prevention, climatic moderation, and a host of other functions whose value cannot be measured. Thus, threats to biodiversity represent a threat to many of the tangible and intangible benefits that other species provide to humans. Much of the attention on biodiversity has focused on tropical rain forests, which have the greatest species diversity of any place on earth. According to United Nations estimates, however, the United States is also one of the richest countries in the world in terms of its biodiversity, ranking eighth in the world in the number of plant species (Groombridge 1992). Patterns of Species Diversity For many years, scientists have been trying to understand differences in species diversity in different places and at different times. We know that species diversity tends to be high in tropical areas, and that it diminishes toward the poles. We know that diversity tends to decrease at higher elevations. Moister climates tend to support more species than drier climates. Areas of greater topographic or environmental variability tend to have more species than more uniform areas (Ricklefs and Schluter 1993; Huston 1994; Rosenzweig 1995). One of the most important trends in biodiversity is the increase in the number of species as the land area increases. Botanists call this trend the species-area relationship, and it is one of the most dependable rules in biology (Connor and McCoy 1979). Simply stated, large areas have more species than small areas. As a rule of thumb, the number of species approximately doubles when the land area is doubled five times (which is the same as multiplying the area by 32). Species diversity can also change over time, and since their arrival, humans have profoundly influenced plant life throughout North America (Crosby 1986). Obviously, a corn field or a parking lot has fewer species than a prairie or forest. On a broad scale, humans have caused the decline, and even extinction, of many species. Thus, we might expect to find lower biodiversity in areas where human land-use is most intense. Many human activities such as construction, urban development, and agriculture influence species directly by disrupting their habitat, by removing other organisms upon which they may depend, or by injuring or killing them. Less direct, but in many cases equally as disruptive to the native species of an area, is the introduction of exotic species. Exotic Species: The Good, the Bad, and the Ugly Exotic species are species that live and reproduce in places where they previously did not occur. The spread of exotic species may occur by natural means, but usually exotic species are spread by humans, either intentionally or unintentionally. Humans introduce exotic plant species for many reasons. Exotic species have been planted as ornamentals, for food crops, and for wind breaks and erosion control. In many cases, humans have spread exotic species unwittingly, for example as weeds associated with grain or livestock. Humans have introduced exotic species to every part of the world. Some exotic species have become a common element of the local biota without causing major ecological consequences. But in many cases, exotic species have threatened the native biota, disrupted ecosystem function, and even wiped out entire populations of native species (Drake et al. 1989). The Good Many of our common agricultural crops, because they originated from areas outside the United States, are exotic species. Wheat (Triticum) and apples (Malus) from the Middle East, rice (Oryza) from Asia, and tomatoes (Lycopersicon) from Central America are only a few examples of crop plants introduced to North America from other parts of the world (Harlan 1976). The Bad Unfortunately, many exotic species have had detrimental effects for both humans and ecosystems worldwide. In parts of the United States, exotic plant species threaten the survival of native species and disrupt ecological processes at multiple levels. For example, the tamarisk (Tamarix), introduced from Asia for erosion control, has become so successful that it has reduced the flow of many rivers in the West, disrupting water supplies and upsetting habitat for many aquatic animals. Aggressive exotic plants may also outcompete native plant species. The Chinese tallowtree (Sapium sebiferum) is rapidly taking over many of the native ecosystems in southeastern Texas, and exotic grass species--many resistant to grazing--have almost entirely replaced formerly dominant native species in California grasslands. The Ugly Still other exotic plant species not only disrupt the local ecology but also act as pests, nuisances, and eyesores. Kudzu (Pueraria lobata), introduced as ground cover in the southeastern United States, spreads rapidly in full sunlight to form a nearly continuous blanket of foliage over the ground layer and trees (Fig. 4-1). The vines choke out other plants, leaving a large area of barren and grotesque trailing sticks when they lose their leaves in the winter.
 Fig. 4-1. A field dominated by kudzu (Pueraria lobata) in northern Georgia. Kudzu, an exotic species native to Japan, was once planted widely for erosion control, but it has become an eyesore and a pest over large areas of the southeastern United States. Botanists know a great deal about these and some other exotic plant species. In many cases, the spread of exotic species is closely related to changes in land use. Many common lawn and garden weeds, such as dandelion (Taraxacum), white clover (Trifolium repens), English plantain (Plantago lanceolata), and timothy grass (Phleum pratense), were introduced from Europe, and their growth is favored over native species by the agricultural practices that Europeans brought to North America.
In this report, we have used information gathered by botanists over the past two centuries to assess general patterns of plant species diversity in North America and to show how these patterns have changed over time. Furthermore, we have tried to assess patterns in the prevalence of exotic species (i.e., species that originated outside North America) and to find out where and to what degree they have become more or less prevalent over time. Our analysis focuses on the number of plant species and the proportion of exotic species at two different spatial scales and at two different points in time for North America. We include estimates for areas covering 1,000,000 ha and areas of 1,000 ha. For perspective, 1,000 ha equals 3.86 mi2, and 1,000,000 ha equals 3,861 mi2. For each area size, we estimate values for the years 1900 and 1996. Thus, our analysis results in four maps of North America for the size of the flora and four maps for the proportion of exotic species. Because our analysis is continuing, and because we must rely on information collected by hundreds of different scientists over more than 200 years, the results are somewhat speculative.
Botanists have been collecting data on the distribution of plants in North America for centuries. Surprisingly, however, we still have much to learn about geographical variations in plant biodiversity and how it has changed over time. Information has been compiled and summarized in many different ways, including plant distribution maps, lists of herbarium specimens, and floras. In this report, we used a large collection of floras. Sources of Data A flora is a list of plant species that are known to occur within a region of interest. Often, the list is accompanied by a scientific description of each species that would permit it to be identified. Botanists have written floras for every type of region imaginable: city, state, and national parks, counties and states, a single pond or rock outcrop, and even for an entire country. Thus, floras span a wide range of land areas, from 1 ha or less to many millions of hectares. Equally varied are the reasons for writing a flora; these reasons include assisting in environmental impact assessment, determining conservation value of a potential nature reserve or park, and simply preparing an authoritative guide to the plant species of a region. Despite the varied types and purposes of floras, we believe they offer great potential for research. Our study represents the first attempt to glean information from thousands of floras spanning the entire continent of North America. We used many different strategies to acquire the floras used in this study. We searched university and government libraries, used computerized index searches, scanned more than 30 key botanical journals, read the bibliography and reference lists of botanical surveys and floras, used government document searches, corresponded with other botanists, and even posted e-mail requests on relevant bulletin boards. Still, we are constantly discovering new floras. For each flora, we determined the total number of plant species and the number of exotic species known to occur within the region covered by the flora. We then ascertained, from the flora itself or from other sources, the latitude, longitude, area, average elevation, and the range of elevations for the region covered by the flora. We also enumerated the number of plant families and genera, which we will analyze in future work. While the number of species in a defined region may be called that region's species richness, we prefer instead to call our data flora size, since the number of species listed depends not only on the number of species actually present but also on the accessibility of the region to botanists and to the history of botanical exploration. In many cases, the author did not specify which species were exotic, so we have begun to determine that ourselves. In this analysis, we used 1,343 floras with data on flora size and 547 floras with data on exotic species (Fig. 4-2).
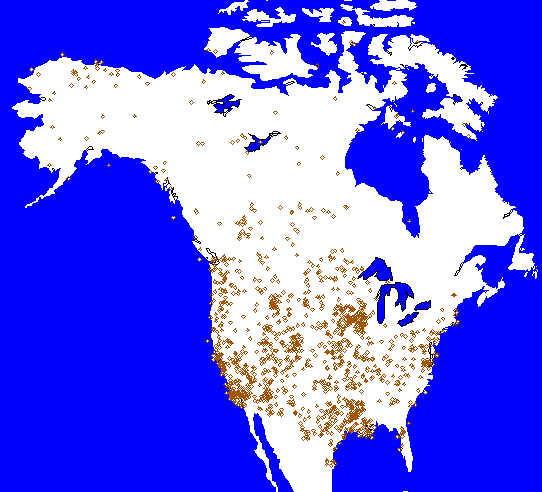 Fig. 4-2. Locations of floras used in this study. Points are centered on the geographic midpoint of each flora. Methods of Estimation Linear regression is a numerical technique used by statisticians and scientists to study relationships between different phenomena. For example, political pollsters use regression techniques to develop relationships between the way people vote and their income, education, and residence location. They then use this information to produce polling results and ultimately to predict the outcome of political races. Similarly, we used the relationships between the numbered of plant species in published floras and some of the characteristics of the areas for which the floras were compiled to estimate the size of the flora for different locations and different times. These characteristics include the area, latitude, longitude, and elevation of the region; the year the flora was compiled; and a measure of the time and effort expended by the author(s) on botanical exploration. We developed mathematical relationships between these characteristics of an area and the size of its flora, and used those relationships to produce the flora maps presented herein. Contour Mapping We used contour maps to illustrate geographic patterns in flora size and proportion of exotic species in North America for two area sizes at two different times. A contour map is used to display geographic patterns of numerical data. The contour lines connect points of equal value, such that two locations lying along the same line have the same value. The contour interval represents the difference in value between adjacent contour lines. Notice that the value for places located between contour lines can only be estimated as some value intermediate between the values of the two contour lines. Regions between contour lines can be shaded using different colors to accentuate the variation. Weather maps commonly use such color schemes to show regions with similar temperatures and rainfall. The placement of contour lines on a map is based on data at known, or estimated, points. Mathematical relationships, in some cases related to regression lines, are used to further estimate values for locations between known points. Generally, accuracy improves as the number of data points increases. Accuracy is relatively high for areas on the map with a high concentration of data points and low where data points are more sparsely distributed (see Fig. 4-2). Sources of Error Although floras hold great promise for biodiversity research, they do have characteristics that may present limitations (Miller and Wiegert 1989; Mayr 1992; Palmer 1995; Palmer et al. 1995). Perhaps most important, no flora is 100% complete. There are almost certainly plants living in any given area that have not been identified and described. In addition, comparisons of floras are difficult because some are more complete than others. Floras have been compiled by thousands of different researchers who all used different methods and who worked to different standards. Some floras were written over many years by professional botanists who spent a lifetime studying one particular region; others were written after only 1 year or a few years of study. Floras also differ greatly in the area covered, from less than 1 ha to millions of hectares. This variety creates difficulties in interpreting differences in diversity, although we have attempted to control for this problem statistically. In the same way, different regions have had different levels of botanical exploration (Fig. 4-2). Other shortcomings of floras include inconsistency in reporting, imprecise definition of study area or collection methods, inconsistency in the definitions of species, and the use of different nomenclature (Kartesz 1994). Thus, variation in reported numbers of species may represent differences in definition rather than differences in biological variability. However, we suspect such problems related to synonymy are relatively minor. Additional sources of uncertainty in these maps include the following:
Despite these shortcomings, we believe floras represent one of the most comprehensive sources of data for plant biodiversity. With due consideration of these limitations and reasonable caution in assessing the accuracy of the results, we believe they provide useful information that is otherwise unavailable.
Size of Flora for 1,000,000 Ha: 1900 and 1996 We estimated the size of the flora for areas of 1,000,000 ha for both 1900 and 1996 (Fig 4-3). As is common for almost all groups of organisms, diversity decreases at higher latitudes. Biologists call this the latitudinal gradient in species diversity. Diversity reaches a maximum in the southeastern United States, which is well known by botanists for its high biodiversity (White and Miller 1988). This rich biodiversity may be due at least in part to the warm, moist climate and relatively mild winters, which are commonly linked to biodiversity. The marked variation in habitat types over moderately sized areas in the southern Appalachians also contributes to this high diversity. Size of flora decreases toward the west and north. Westward decreases may be related to the increasingly dry climate, less environmental variation, and the relatively shorter history of exploration. With its long settlement history, the flora of eastern North America is more comprehensively known than that of the West. Note that the size of the flora has increased in 1996 relative to 1900, especially in the mountain West. The increase in size of flora over time is probably not due to biology alone. Development in the West has provided increased accessibility, and the flora has probably become more thoroughly described throughout the continent since the turn of the century. Nevertheless, greater diversity in the Rocky Mountain region compared to the surrounding areas is probably real and may be attributed to the wide range of elevation, and thus the high diversity of potential environments, in this mountainous region. Size of Flora for 1,000 Ha: 1900 and 1996 The flora size for areas of 1,000 ha for both time periods is much smaller than for the 1,000,000-ha scale, simply reflecting a smaller area (Fig. 4-3). The decrease in diversity from east to west remains, although the peak in the Southeast is less pronounced. In addition, the latitudinal gradient is reduced. The slightly enhanced diversity in Alaska may reflect the relatively high diversity of some coastal, temperate rainforest environments. These environments harbored many species adapted to cool climates when glaciers covered much of the interior during Pleistocene ice ages.

From 1900 to 1996, flora size increased for much of the West but decreased in parts of the East. The increase in the West probably reflects increased knowledge and better documentation of the floras. The possibility that floras are decreasing in size in eastern North America is troubling and could possibly be a result of continued human population growth, development, and urban encroachment. However, we believe it is more likely that the flora size has decreased in the East due to growing interest among botanists in preparing floras for very unique environments such as rock outcrops, bogs, and ponds. Although these environments often house rare and unique species, they are typically depauperate; that is, they harbor relatively few species. Thus, increased focus on these special environments would lead to lower average flora size for small areas, but not, as observed above, for larger areas, which already incorporate these special habitats. More data from floras in the East are needed to better understand this pattern.
We determined the proportion (in percent) of the flora that consists of exotic species for an area of 1,000,000 ha in 1900 and 1996 (Fig.4-4). The resulting maps show the proportion of exotic species. For a region covering 1,000,000 ha, the proportion of exotic species is greatest in the mid-Atlantic region and New England. This high percentage of exotic species probably reflects the very high human population density, the intensity of development and disturbance, and the long history of European settlement there. The proportion of exotic species also increases near the Pacific Coast, probably due to proximity to settlement and development and a long history of European settlement, initially by the Spanish. The proportion of exotic species near both coasts has increased since 1900. The spread of exotics from port cities is not surprising, because it is likely that seeds were inadvertently introduced in ballast, packing materials, and soil arriving on ships at these ports. There is a pronounced minimum of exotic species toward the middle of the continent, reflecting the shorter development history and greater isolation, which would tend to lower the probability of invasion by exotic species. In 1996, however, there appears to be a trend of increasing exotic species bulging northward from Mexico and the Caribbean into the southern plains. There is also a pronounced latitudinal gradient in proportion of exotic species. There are three possible reasons why fewer exotic species occur in the northern latitudes: (1) relatively few exotic species can survive the extreme cold, (2) most species adapted to extreme cold already occur throughout the Arctic, and (3) there has been relatively little human influence because human population density is low and consequently landscapes are relatively undisturbed.
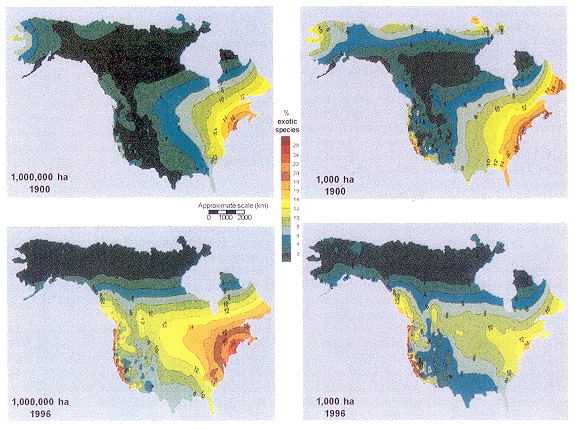 Fig. 4-4. Estimated proportion (percentage) of the flora consisting of exotic species for areas of 1,000 and 1,000,000 ha in 1900 and 1996. The maps can be interpreted by considering any point on the map as the center of an area of the specified size. An area of 1,000,000 ha is equivalent to the area enclosed by a circle with a diameter of about 70 mi; 1,000 ha is equivalent to the area enclosed by a circle with a diameter of slightly more than 1 mi.
The same patterns noted for 1,000,000 ha appear to hold for the smaller 1,000-ha area (Fig. 4-4), although the values are much higher for the smaller areas. This increase occurs because many exotic species are widespread and tend to occur repeatedly in numerous floras. Although exotic species proportions appear to be increasing in most places, the proportion of exotics in 1,000 ha appears to be declining in the Southeast. This finding probably reflects botanists' increasing tendency to seek out undisturbed, intact natural areas, preserves, and unique habitats for floral investigation. These intact habitats typically support few exotic species, and their increased representation among floras would lead to a reduction in the average level of exotic species.
In this project we have demonstrated the potential value of using the data contained in floras to study geographical patterns of plant species diversity and frequency of exotic species. Despite their limitations, floras contain valuable and comprehensive information on the plant life of North America that may not exist elsewhere. Moreover, by tracing floristic data collection through history, we can ascertain long-term trends and changes in these patterns over time. Our results are consistent with many previously identified trends in diversity (Ricklefs and Schluter 1993; Huston 1994; Rosenzweig 1995). Flora size increases as the size of the study area increases. Diversity decreases toward higher latitudes, especially over larger study areas. Flora size in North America is greatest in the southeastern United States. The high diversity may be related to the region's warm, humid climate, which is thought to be favorable for plant growth (Currie and Paquin 1987). Flora size in North America decreases toward the west, and this decrease may reflect, in part, drier environments as well as a less comprehensively documented flora. By investigating floras compiled over a long period of time, we can assess changes over time in the size of a flora and the percentage of exotic species and relate these changes to historical patterns of land use. Increases in the size of floras since 1900 probably do not reflect increased plant biodiversity but rather mirror settlement patterns and land-use changes. As the West has been more thoroughly settled over the past 100 years, the plant life has been more thoroughly documented and the size of floras has therefore increased. The reason for the decreased size of floras in the eastern United States since 1900 is unclear, but it may be a result of increased botanical interest in unusual environments that house relatively few, but unique, plant species. Repeated inventory of plants in areas that were previously studied intensively may shed an interesting light on this question. The patterns of exotic species distributions appear more directly and more profoundly linked to land use and land-use history. Proportions of exotic species are greatest in the northeastern United States and the Atlantic coast of Canada, which have the longest continuous history of European settlement and the highest population density in North America. The west coast also has relatively high proportions of exotic species, and it too has a long history of development dating to the eighteenth century, a large population, and intensive development over large areas. It is noteworthy that both of these regions are home to North America's largest ports and that both coasts have served as a gateway to tens of millions of human immigrants. Undoubtedly, along with the humans came many species of exotic plants. The proportion of exotic species is lowest in the mountain West and in the far northern latitudes of Canada and Alaska, which are sparsely populated, minimally developed, and relatively isolated from sources of exotic species. Moreover, these cold, harsh environments require special adaptations that few exotic species have acquired. This project, though preliminary in nature, has revealed distinct insight into the geographic variation in plant species diversity and the prevalence of exotic species in North America. Collection of additional data from floras in areas that are poorly represented at present (Fig. 4-2) will lend greater reliability and may shed additional light on the patterns discussed here. The resulting understanding of trends in the geographical distribution of biological diversity will be essential in developing strategies to best conserve it and to benefit from it.
The Baltimore-Washington Regional Collaboratory Land-Use History Research Program
Text-only |